|
|
|
|
|
|
De Cel.Cel : ( Lat. cellula , verkleinwoord van cella =
kamer ) de meestal microscopisch kleine structurele eenheid waarvan elk
organisme er één of meer bevat. 1. GESCHIEDENISDe 17de-eeuwse Engelse onderzoeker
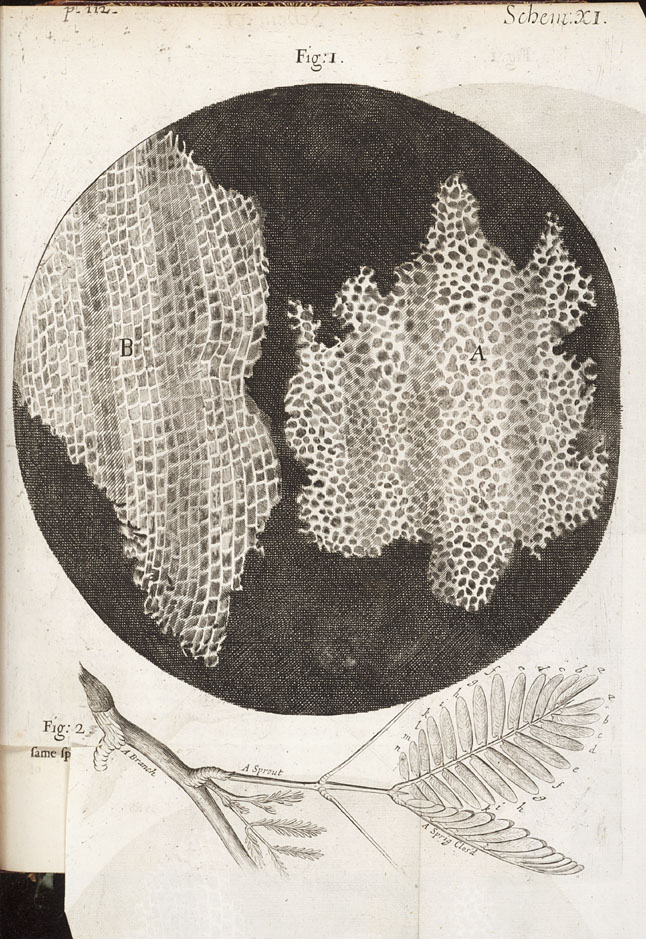
Robert Hooke bestudeerde onder de microscoop dunne plakjes flessenkurk.
Het materiaal bleek te zijn opgebouwd uit kamertjes, enigszins
vergelijkbaar met de cellen van een honingraat. Hij noemde ze (1667)
cellulae. In volgende jaren werden overeenkomstige structuren bij
ander plantenmateriaal beschreven. Bij levend materiaal waren de cellen
gevuld met wat beschreven werd als een 'slijmerige vloeistof'. In 1831 zag
Robert Brown hierin een lichaampje, de celkern. Ook bij dierlijk
materiaal werden beide componenten, 'slijmerige vloeistof' en celkern,
gevonden. Dit leidde tot het inzicht dat alle organismen uit deze twee
eenheden zijn opgebouwd (de celtheorie van Schleiden en Schwann,
1839). Voortgezet onderzoek leerde dat de 'slijmerige vloeistof' de drager
is van de levensverschijnselen; zij kreeg de naam protoplasma (Purkinje
1840 voor dieren, von Mohl 1848 voor planten). Tegenwoordig spreekt men
van cytoplasma en wordt de term protoplasma gebruikt voor
cytoplasma plus kern. Met ectoplasma wordt bedoeld het meest naar buiten
gelegen cytoplasma. De wereld van de cel is voor het blote oog onzichtbaar. Zij bleef dan ook volledig onbekend en onontdekt tot halverwege de zeventiende eeuw, toen mensen met een onderzoekende geest, geholpen door vaardige handen, lenzen begonnen te slijpen en deze gebruiken om hun gezichtsvermogen te vergroten. Een van de eerste microscoopbouwers was de Engelsman Robert Hooke – Fysicus , meteoroloog, bioloog, ingenieur, architect – een zeer bijzonder kind van zijn tijd. In 1665 publiceerde hij een schitterende verzameling tekeningen, Micrographia getiteld, waarin hij zijn microscopische waarnemingen beschreef. Daarbij bevond zich ook een tekening van een dun schijfje kurk dat een honingraatstructuur vertoonde, een regelmatig netwerk van ‘microscopisch kleine poriën’ of ‘cellen’.
Een van Hooke’s meest begaafde tijdgenoten was de Nederlander Antonie van Leeuwenhoek, die meer dan tweehonderd microscopen van een speciaal type gemaakt heeft. Zij bestonden eenvoudig uit een klein glazen bolletje, vastgekit in een koperen plaat. Door dit instrument dicht bij zijn oog te houden en door de glazen knikker te kijken naar voorwerpen, vastgeprikt op een naald die hij met behulp van een schroef kon manipuleren, slaagde Van Leeuwenhoek erin om voorwerpen tot 270 maal te vergroten en deed hij zeer opmerkelijke ontdekkingen. Hij kon voor het eerst in bloed, sperma en in het water uit plassen en sloten dagene zien wat hij zelf ‘animalcules’ noemde. Zelfs zag hij bacteriën, die hij zó nauwkeurig tekende, dat specialisten ze vandaag de dag nóg kunnen determineren.
Antonie van Leeuwenhoek
In 1827 slaagde de Italiaanse fysicus Giovanni Battista Amici erin om de belangrijkste optische vertekeningen van lenzen te corrigeren. De beelden werden op slag reusachtig veel scherper; zoveel, dat men slechts enkele jaren later al de algemene theorie kon formuleren dat alle planten en dieren uit één of meer gelijke eenheden, de cellen, zijn opgebouwd.
Giovanni Battista Amici
Rond 1900 was een aanzienlijk aantal celbestanddelen beschreven en van een naam voorzien. De onderzoekers raakten echter steeds meer verstrikt in een nieuw probleem, dat onoplosbaar scheen omdat het gevormd werd door de wetten van de natuurkunde zelf. Zelfs met een volmaakt instrument kan men geen details weergeven die kleiner zijn dan ongeveer de halve golflengte van het gebruikte licht, en dat betekent dat het oplossende vermogen van een gewone lichtmicroscoop bij ongeveer 0.25 µm ligt. De doorbraak van 1945: De langdurige, ononderbroken inspanning begon aan het eind van de Tweede Wereldoorlog plotseling vruchten af te werpen, toen als gevolg van een merkwaardige samenloop van omstandigheden, een veelheid aan waardevolle nieuwe instrumenten en techniek min of meer tegelijkertijd ter beschikking kwam. De morfologen werden verblijd met de elektronenmicroscoop. Dit instrument, dat in de jaren 30 werd uitgevonden, heeft een oplossend vermogen dat groot genoeg is om het gehele, onbekende celgebied, tot op de nanometer, te doorvorsen. In verbazingwekkende korte tijd slaagde men erin methoden te ontwikkelen om objecten te prepareren en construeerden ze instrumenten die de objecten in ultradunne plakjes sneden. Men verkreeg steeds betere beelden, zodat al in het begin van de jaren zestig een groot deel van het onbekende gebied morfologisch in kaart gebracht werd.
2. HOOFDTYPENCellen die eenvoudig van bouw zijn en geen echte kern bezitten, worden PROKARYOOT genoemd. Het DNA zit weliswaar geconcentreerd in één gebied in de cel (de nucleoide), maar dat gebied wordt niet door een membraan afgescheiden van de rest van de cel. Men treft ze aan bij de bacteriën (inclusief de cyanobacteriën) en archaea. Het onderzoek van de prokaryote cellen vormt het gebied van de microbiologie. Alle overige organismen
(eencellige en meercellige) bezitten cellen met een kern en een
ingewikkelder bouw. Deze cellen worden eukaryoot genoemd. Het
onderzoek van de eukaryote cel wordt beoefend in de celbiologie. Ligt in
een meercellig organisme een groot aantal gelijkvormige cellen bijeen, dan
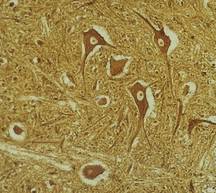
spreekt men van een weefsel . De studie van het verband tussen de cellen
van een meercellig organisme wordt tot de weefselleer of histologie
gerekend. 3. BOUWProkaryote cellen zijn eenvoudig gebouwd. Zij worden beschreven bij de bacteriën en Cyanobacteriën. Hierna worden uitsluitend
de eukaryote cellen behandeld. 3.1 GrootteDe grootte van cellen is zeer variabel; de kleinste dierlijke cellen zijn ongeveer 4 µm in diameter, de grootste – bijv. de dooier van een vogelei – enige centimeters. In geïsoleerde toestand zijn dierlijke cellen gewoonlijk bolrond of spoelvormig; in weefselverband zijn zij plat prismatisch of langgerekt prismatisch of kubisch; door wederzijdse samendrukking worden het veelvlakken. Sommige cellen hebben geen vaste vorm, maar zijn variabel, zoals de amoeboïde cellen uit het bloed.
De cellen van planten lopen in vorm en
grootte sterk uiteen. Gemiddelde afmetingen zijn 15 – 100 µm, maar
plantenvezels kunnen zeer lang zijn, bijv. van katoen, hennep en vlas tot
50 mm. Bij enkele zeewieren (Valoniaceae) bereiken de cellen een inhoud
van 10 ml. Men vindt bij de hogere planten bolronde, cilindrische, kubus
-, plaat - of vezelvormige cellen. Ook de eencelligen vertonen een grote
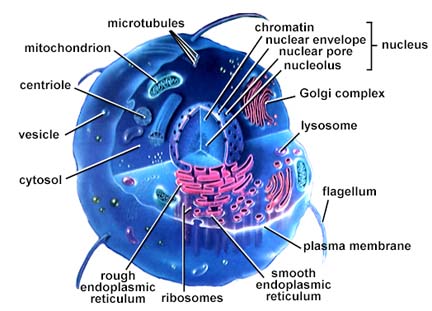
vormverscheidenheid. 3.2 MembranenDe cellen van eukaryote
organismen bevatten verschillende typen membranen. Tezamen worden deze
biomembranen genoemd, dit in tegenstelling tot kunstmatige membranen .Eukaryote cellen bestaan
steeds uit een door een celmembraan omgeven hoeveelheid cytoplasma,
waar een aantal celorganen (organellen) van verschillende vorm,
grootte en functie in zijn ingebed. Alle organellen (behalve de ribosomen)
zijn omgeven door een membraan: intracellulaire membraan. Bij
sommige organellen is dit membraan in feite een dubbele membraan, hetgeen
een aanwijzing is dat dit organel vroeger zelfstandig leefde en is
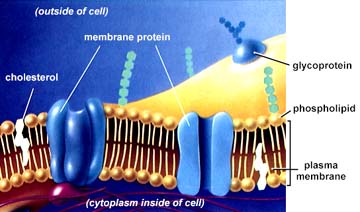
opgenomen door de eukaryote cel (zie endosymbiose). 3.2.1 BouwIn beginsel hebben al deze membranen dezelfde bouw. Alle membranen bestaan uit lipiden en eiwitten. De lipiden zijn voornamelijk fosfolipiden, hoewel in sommige membranen ook cholesterol een belangrijk bestanddeel vormt. De fosfolipiden vormen een dubbellaag met een dikte van 8–10 nanometer waarbij hun hydrofiele groepen naar buiten zijn gekeerd terwijl de hydrofobe vetzuurstaarten naar elkaar toe zijn gericht. Deze dubbellaagstructuur is met behulp van de elektronenmicroscoop goed waarneembaar. De eiwitten die deel uitmaken van het membraan worden onderverdeeld in twee groepen. De intrinsieke of integrale eiwitten bevinden zich geheel of gedeeltelijk in de fosfolipidedubbellaag. Sommige van deze eiwitten steken zelfs dwars door de dubbellaag heen. In dat geval spreekt men van een transmembraaneiwit. Tot voor kort werden de eiwitten in de membraan beschouwd als een vrij homogene groep; dit is echter maar zeer ten dele het geval. Er komen veel geconjugeerde eiwitten voor, in het bijzonder eiwitten die geassocieerd zijn met koolhydraten (glycoproteïnen) en in mindere mate met lipiden (glycolipiden). Koolhydraatketens spelen een belangrijke rol bij de communicatie tussen verschillende cellen (herkenning, hechting). De laag koolhydraatgroepen speelt een rol bij de totstandkoming van de celmantel (glycocalyx) aan de buitenzijde van de celmembraan. Zowel de fosfolipidemoleculen als de eiwitten bezitten een zekere beweeglijkheid in het vlak van het membraan (laterale diffusie). Er vindt daarbij echter geen uitwisseling van moleculen tussen de beide helften van de dubbellaag plaats. Ook de oriëntatie van de eiwitmoleculen ten opzichte van beide helften van de dubbellaag blijft hetzelfde. De beide zijden van het membraan zijn dus niet identiek, iets wat direct samenhangt met de biologische functie van biomembranen. De mate van vloeibaarheid is afhankelijk van de samenstelling van het membraan. De aanwezigheid van bijv. onverzadigde vetzuurketens in de fosfolipiden verhoogt deze vloeibaarheid. De aanwezigheid van cholesterol daarentegen verlaagt de beweeglijkheid van de moleculen in de dubbellaag. Ook bij de sterkste vergroting zijn de celmembranen van twee naburige cellen altijd van elkaar gescheiden door een tussenruimte van ten minste enkele tientallen nm, terwijl niet altijd duidelijk is of deze ruimte opgevuld is met materiaal. Deze afstand berust waarschijnlijk mede op een patroon van kleine elektrische ladingen aan de buitenzijde van het celoppervlak. Waarschijnlijk wordt niet alleen door elektrostatische afstoting van gelijknamige ladingen verhinderd dat de cellen elkaar raken, maar kan ook door aantrekking van ongelijknamige ladingen een zeer specifieke onderlinge attractie tussen cellen (cellulaire adhesie) tot stand komen, die oorzaak is voor de inwendige samenhang van weefsels. Vaak echter is de afstand tussen de cellen veel groter dan enkele tientallen nm en gevuld met intercellulaire substanties. Deze hebben vaak een polysacharide karakter (mucopolysacharide, chitine), terwijl ook eiwitvezels kunnen voorkomen (bijv. collageen) of andere specifieke verbindingen (calciumfosfaat). In de celmembraan van gespecialiseerde cellen kan men aparte structuren vinden. Cellen van het dunne-darmslijmvlies bijv., die vooral als functie hebben het opnemen van voedingsstoffen uit de darmholte, vertonen een groot aantal uitstulpingen van de celmembraan (microvilli), waardoor het vrij aan de darmholte gelegen celoppervlak zeer sterk vergroot wordt. Lichtmicroscopisch is deze structuur waarneembaar als borstelzoom. Opperhuidcellen (zie huid), die onderling een sterke samenhang moeten vertonen, bezitten vaak desmosomen, verdikkingen die brugvormige verbindingen vormen met dergelijke verdikkingen op andere celmembranen. De betekenis van de membranen is tweeledig, aan de ene kant die van een scheiding tussen de cel (het organel) en de omgeving, aan de andere kant juist die van een verbinding tussen beide: alle stoffen die de cel of het organel in - of uitgaan moeten de membraan passeren. De membranen zijn selectief permeabel, dwz. zij laten sommige stoffen gemakkelijk, andere moeilijk tot vrijwel niet door.
3.2.2 FunctieBepaalde in water oplosbare stoffen (bijv. ionen) kunnen de membraan ongehinderd passeren (passief transport). Men neemt aan dat de transmembraaneiwitten, eiwitten die door de hele membraan steken, een passageplaats vormen voor ionen, die uitgewisseld kunnen worden tussen het extracellulaire en het intracellulaire compartiment. Deze eiwitkanalen corresponderen met de membraanporiën waarvan het bestaan reeds door celfysiologen was verondersteld. Voor een groot aantal stoffen vormt de membraan een barrière, zodat aan weerszijden van het membraan heel verschillende condities heersen, bijv. wat betreft de concentratie van ionen en van producten van het cellulaire metabolisme. Om deze verschillen te kunnen handhaven, beschikken membranen over een groot aantal specifieke transportsystemen, elk bestaande uit één of meer eiwitten, die in staat zijn stoffen tegen een concentratiegradiënt in door het membraan heen te transporteren. Voor dit actieve transport is energie nodig, meestal in de vorm van ATP. Op deze manier kunnen bijv. voedingsstoffen zoals glucose, die in het milieu van de cel vaak in kleine concentratie aanwezig zijn, binnen de cel geconcentreerd worden. Een ander bekend voorbeeld is de handhaving, ongeacht de omstandigheden buiten de cel, van een lage concentratie natriumionen (Na+) en een hoge concentratie kaliumionen (K+) in de cel door de natrium-kalium pomp in het plasmamembraan. Deze pomp speelt ook een essentiële rol in de geleiding van zenuwimpulsen (zie impuls [fysiologie]). Verder is de energieproductie in de eukaryote cel voor een belangrijk deel afhankelijk van het handhaven van een verschil in de concentratie van de waterstofionen (H+) ter weerszijden van de membranen die de mitochondriën omgeven. Ook zijn membranen betrokken bij de regulering van de cellulaire activiteit door hormonen. De voor opname van stoffen uit het extracellulaire milieu zo belangrijke receptoren (waaraan een specifiek op de receptor passende ligand zich kan binden) zijn veelal ook eiwitten. Bij de reeds aangeduide transporten, actief dan wel passief, gaat het om stoffen met betrekkelijk kleine moleculen, zoals ionen, enkelvoudige suikers (monosachariden). Grotere moleculen zoals bijv. eiwitten en polysachariden kunnen de celmembraan niet passeren, maar kunnen door vorming en fusie van door membranen omgeven blaasjes naar binnen of naar buiten worden gesluisd. Het proces waarbij materiaal (bijv. een secretieproduct) dat in een blaasje is ingesloten naar buiten wordt gebracht heet exocytose. Wanneer materiaal wordt
opgenomen doordat de celmembraan instulpt en een blaasje vormt, spreekt
men van endocytose. Gaat het hierbij om zekere hoeveelheden
vloeistof, dan spreekt men van pinocytose; betreft het
microscopisch waarneembare deeltjes, bijv. bacteriën, dan spreekt men van
fagocytose. De membranen om de organellen brengen mee dat de cel
verdeeld is in compartimentjes waarbinnen zich biochemische reacties
kunnen voltrekken zonder stoornis door erbuiten aanwezige stoffen. Zo zijn
de enzymen die de opbouw van een stof verzorgen in andere compartimenten
aanwezig dan de enzymen die de afbraak van dezelfde stof verzorgen. De
selectieve permeabiliteit van de membranen is aan het leven gebonden, ze
verdwijnt bij afsterven van de cel, wat leidt tot afbraak van de
celbestanddelen door de eigen enzymen: autolyse van de celinhoud. 3.3 CytoplasmaHet cytoplasma bevat eiwitten die deels een enzymfunctie hebben; het heeft in actieve cellen een hoog watergehalte (60–95%, in plantaardige weefsels soms tot 98%). In het water zijn ionen en tal van laagmoleculaire organische stoffen opgelost, zoals aminozuren en suikers. De zijketens van de
aminozuren, die de polypeptideketens van de eiwitten opbouwen, zijn zeer
verschillend van aard (basisch, zuur, neutraal, wateraantrekkend of
-afstotend); zij kunnen gemakkelijk met andere zijketens reacties aangaan
en zo bruggen vormen tussen naburige eiwitmoleculen. Sommige bruggen zijn
zeer gevoelig voor de temperatuur, andere voor de plaatselijke zuurgraad (pH)
of oxidatietoestand (redoxpotentiaal); kleine veranderingen in deze
grootheden kunnen de vorming of het verbreken van die bruggen sterk
beïnvloeden. Deze als hechtpuntentheorie bekend staande
voorstelling verklaart zowel de bovengenoemde eigenschappen van het
cytoplasma als de grote veranderingen die op korte termijn in zijn
viscositeit kunnen optreden. In het cytoplasma vinden o.a. de afbraak van
glucose tot pyrodruivenzuur (glycolyse) en de activering van aminozuren
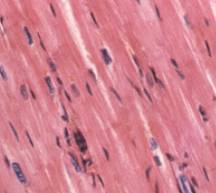
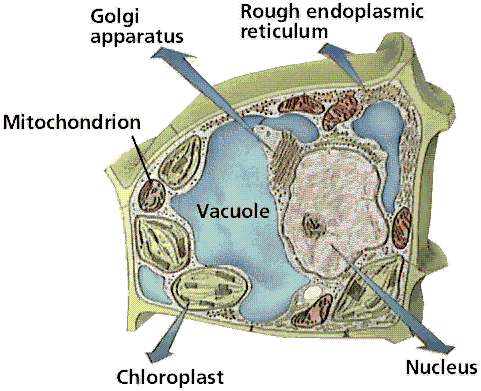
plaats. 3.4 OrganellenHet cytoplasma is doortrokken door een netwerk van vertakte buisjes, die door een 'unit membrane' zijn omgeven, het endoplasmatisch reticulum (= endoplasmatisch netwerk, veelal afgekort als ER). Deze buisjes staan in verbinding met de kernmembraan en zijn van belang voor het transport van stoffen binnen de cel. Men onderscheidt glad en ruw ER. Het ruwe is bezet met een groot aantal ribosomen, die bij het gladde ontbreken. Het ruw ER vervoert de eiwitten die geassembleerd worden op de ribosomen, en bedoeld zijn om door de cel uitgescheiden te worden. De polypeptideketens glijden door de poriën in de wand van het ER en komen terecht in de ruimte omgeven door het membraan (de cisternale ruimte). Vaak wordt in de cisternale ruimte een koolhydraat aan de eiwitten toegevoegd, zodat een glycoproteïne gevormd wordt. Daarna omwikkelt het ER de uit te scheiden stoffen met een membraan, zodat een blaasje (Eng.: vesicle) ontstaat. Deze blaasjes transporteren de glycoproteïnen door het cytoplasma richting celmembraan. Naast haar functie in de secretie van produkten, staat het ruw endoplasmatisch reticulum ook in voor de aanmaak van membranen. Deze kunnen aan het bestaande ER vastgehecht blijven, of naar andere delen van de cel bewegen onder de vorm van transportvesicles. Het gladde ER staat vaak in verbinding met het ruwe ER. Toch betreft het een geheel ander organelcomplex dat in de meeste cellen minder op de voorgrond treedt, maar betrokken kan zijn bij zeer uiteenlopende celfysiologische processen (bijv. de productie van steroïde hormonen in de schors van de bijnier). De ribosomen zijn zeer kleine lichaampjes die uit ribonucleïnezuur (RNA) en eiwit zijn opgebouwd en verantwoordelijk zijn voor de synthese van de eiwitten (zie eiwitvorming). Behalve buitenop het ER bevinden de ribosomen zich ook in het cytoplasma (waar enige samen een polysoom kunnen vormen), in de mitochondriën en bij planten in de plastiden. Het zijn de enige organellen die niet door een membraan zijn omgeven. De eiwitten die in de ribosomen van het ruw ER gemaakt worden, zullen veelal later naar verschillende celorganellen getransporteerd worden, maar kunnen door de cel ook naar buiten gebracht worden (transporteiwitten), bijv. in de vorm van een secretieproduct. De verdeling van deze eiwitten over hun verschillende eindbestemmingen vindt meestal plaats in een tussenstation, het Golgi-apparaat, ook wel dictyosoom genoemd. Dit is een groepje van door een membraan omgeven kleine holtes waarin de synthese van stoffen voltooid wordt. Dit betreft met name het modificeren, sorteren en afleveren van producten die door het ruwe ER zijn aangevoerd. Van het Golgi-apparaat kunnen zich kleine transportblaasjes afsnoeren en zich verplaatsen naar de buitenmembraan, waar zij mee versmelten, onder uitscheiding van de inhoud naar de buitenzijde van de cel. Zo worden door kliercellen (pro-)enzymen en andere klierproducten naar het afvoerkanaal afgescheiden, terwijl bij planten op dezelfde manier bouwstenen voor de celwand te bestemder plaatse belanden. Men heeft vroeger onder de naam microsomen vele kleine lichaampjes samengevat waarvan inmiddels gebleken is dat er enige soorten met verschillende bouw en biochemische functie onderscheiden kunnen worden, zoals lysosomen, die rijk zijn aan hydrolasen (een bepaald type enzymen), die vrij komen bij beschadiging van de cel en dan de afbraak van de eiwitten, nucleïnezuren en andere bestanddelen van de beschadigde cel katalyseren; fagosomen, die ontstaan door opname van hoogmoleculair materiaal van buiten de cel, zelf geen enzymen bezitten, maar door versmelten met lysosomen hydrolyserende enzymen kunnen gaan bevatten (en dan geen fagosomen meer zijn); peroxisomen met een enzymsysteem dat de oxidatie van glycolaat katalyseert; glyoxisomen met een enzymsysteem van belang voor de afbraak van vetten. De laatste bevatten enzymen voor o.a. de glyoxylaatcyclus. In de mitochondriën wordt de in koolhydraten en vetten aanwezige energie overgebracht via de vorming van ATP ter beschikking gesteld van energievragende reacties in de cel. Het aantal van de hiervoor genoemde soorten organellen per cel is zeer verschillend en hangt nauw samen met de activiteitstoestand van de cel. Zo vindt men er in de cellen van rustende plantenzaden weinig en neemt hun aantal bij beginnende kieming sterk toe. De celkern (nucleus of karyon) is het grootste en duidelijkste lichaampje in iedere cel. Meestal is de kern bolrond, soms langgerekt; bij de meeste organismen is er één per cel. De grootte van de celkern ligt veelal tussen ca. 5 en 20 micrometer, zeer kleine kernen worden bij schimmels gevonden (tot 0, 5 µm), grote bij veel vertegenwoordigers van de Leliefamilie (ca. 50 µm), de grootste kernen worden gevonden in de eicellen van enige palmvarens (tot ca. 600 µm). Als regel is er één kern per cel, maar er zijn ook lagere planten waar de verdeling in cellen niet volledig is en waar dan veel kernen in een cel voorkomen; men spreekt dan van coeloblasten. Dit is bijv. het geval bij het zeewier Caulerpa prolifera, waar elke coeloblast uiterlijk overeenkomst vertoont met een gehele hogere plant, met blad-, stam- en wortelachtige delen. De rustende kern is omgeven door een membraan (soms dubbel) waar poriën in voorkomen. De membraan omsluit de kernvloeistof (karyolymfe), waar zich de chromosomen en een of meer kernlichaampjes (nucleoli, enkelv. nucleolus) in bevinden. De chromosomen bestaan uit nucleoproteïden, verbindingen van nucleïnezuren (DNA) en eiwitten. In de nucleïnezuurketens is de erfelijke aanleg van het organisme gecodeerd. De kern is vrijwel de enige plaats waar de nucleïnezuren (DNA zowel als RNA) aangemaakt worden. In de rustende kern zijn de chromosomen meestal niet te zien, maar tijdens de kerndeling (zie celdeling) zijn ze in levende toestand onder de fasecontrastmicroscoop goed waar te nemen; ze zijn het meest bestudeerd in gefixeerde en daarna met basische kleurstoffen gekleurde preparaten. Alles wijst erop dat deze chromosomen ook buiten de periode van de kerndeling (de interfase) intact aanwezig zijn, al zijn ze dan onder het lichtmicroscoop niet zichtbaar. Het aantal chromosomen is normaliter voor ieder soort organisme vast; zie evenwel polyploïd. Trilharen (ciliën), soms ook zweepharen (flagellen), komen op het buitenoppervlak van de cel voor bij eencelligen, sommige wieren, in trilhaarepitheel bij dieren en bij de spermatozoön der dieren en de spermatozoïden van de lagere planten (tot en met de varens en sommige naaktzadigen). Alle vertonen een grote overeenkomst in bouw (zie cilia). Het geheel is omgeven door een uitstulping van de celmembraan.3.5 StevigheidDe cel moet bestand zijn tegen de vele krachten die binnen en buiten de cel plaatsvinden, bijv. die welke veroorzaakt worden door stromingen buiten de cel (bijv. bloed bij vele meercelligen, water bij eencelligen) en door de bewegingen van de organellen in de cel. De cel maakt daartoe gebruik van het principe van tensegrity: de wijze waarop deze krachten worden verdeeld over het celskelet en de celorganellen. Veranderingen in het celskelet - waaraan enzymen zijn vastgehecht - kunnen verantwoordelijk zijn voor het in werking stellen van die enzymen. Mogelijk is de invloed van tensegrity zelfs tot in de celkern aanwezig, waardoor tensegrity kan leiden tot het mechanisch aan- of uitzetten van genen. De organellen dobberen niet zomaar rond in hetcytoplasma, maar zitten ingebed in een netwerk van vezels, het cytoskelet. Het cytoskelet zorgt voor structurele steun en voor de mobiliteit van cellen. Speciale onderdelen, de motormoleculen, veroorzaken de bewegingen van ciliën en flagellen; ze zorgen er ook voor dat spiercellen kunnen samentrekken. De organellen gebruiken het cytoskelet ook als een soort monorail om zich doorheen de cel te verplaatsen. Ciliën en flagellen zijn draadvormige structuren aan de buitenzijde van cellen, die instaan voor de beweging. Ze worden aangetroffen bij vele ééncelligen, maar ook bij zaadcellen van dieren, algen en sommige planten. De ciliën op de cellen van organen dienen om vloeistof over het weefsel te laten vloeien. De cellen die de luchtpijp van de mens aflijnen, bijvoorbeeld, verwijderen zo mucus met afvalpartikeltjes uit de longen. Flagellen en ciliën verschillen onderling in lengte, aantallen per cel en bewegingspatroon. Flagellen zijn 10 tot 200 mm lang, vertonen een golfvormige beweging, en komen gewoonlijk alleen of in kleine aantallen per cel voor. Ciliën zijn 2 tot 20 mm lang, bewegen als roeiriemen, en komen in grote aantallen per cel voor. Beide structuren zitten vol microtubuli, één type vezels dat onderdeel uitmaken van het cytoskelet. Een ander type vezels, microfilamenten of actine filamenten, zorgen voor de samentrekking van spiercellen. Het zijn volle staafjes, opgebouwd uit het eiwit actine. Duizenden van die actine filamentjes liggen parallel aan elkaar in de lengterichting van de spiercel. Tussen elke laag actinemoleculen ligt een laag myosine. De contractie van de spiercel ontstaat doordat de actine en myosinefilamenten over elkaar schuiven.
|
 |
Klik hier voor een printvriendelijke versie van deze pagina |
|
|
Download een Word-versie van deze pagina |
 |
Hoofdmenu |
|
|
|
|
|
|